
Matter That Reads Itself :
The physics of why life interprets rather than merely reacts

A physicist asked how molecules become messages. His answer changes what it means to say “life is interpretation.”
A few months ago I wrote a piece called “Life Is Interpretation.” The claim was straightforward on the surface: biomolecular condensates—those membrane‑less, sticky, self‑assembling compartments now being found everywhere inside cells—don’t just perform chemistry. They read, write, and rewrite the cell’s own state. They embody an interpretive loop, and that loop, I argued, is not a quirk of complex organisms. It is the defining activity of living matter at every scale. The essay closed with a line that was meant to be provocative but also precise: “Life is what happens when matter organizes itself to read.”
What I didn’t do in that piece—and what I want to do here—is show where the intellectual scaffolding for that statement actually comes from. It belongs to a physicist named Howard Pattee, a man who spent more than fifty years asking a question so simple it sounds naive: How does a molecule become a message?

Pattee’s answer is the most rigorous physical account we have of why life is not just chemistry, and why interpretation is not a metaphor you layer on top of biology but an architecture you can locate in the physics itself. My condensates argument, if it holds, is a concrete materialization of Pattee’s logic. But to see why that matters, you have to understand the logic first—and why Pattee himself never called his work “biosemiotics,” even though biosemioticians now claim him as a pillar.
What follows is not a summary. It’s an attempt to walk you through Pattee’s core architecture in plain language, to connect it to the condensate story, and to ask what the whole framework looks like once you combine a physicist’s epistemic cut with the idea that interpretation really is a form of matter.
Why Standard Physics Doesn’t See Life
The problem begins with something most people never notice because their physics textbook taught them to ignore it. Physics speaks in two completely different languages, and it almost never admits that.
The first language is dynamical laws—equations that tell you how a system evolves over time, driven by energy differences. Set the initial conditions, turn the crank, and the future unwinds. Newton’s laws, thermodynamics, Schrödinger’s equation: they all push matter forward. In this language, alternative pathways do not exist except as artifacts of human ignorance.
The second language is constraints. A constraint is not a law. It’s a local, historical structure that says “thus far and no further.” A cell membrane is a constraint. The folded shape of a protein is a constraint. A particular sequence of DNA is a constraint. None of these can be derived from Maxwell’s equations. They are what Murray Gell‑Mann called frozen accidents—arrangements that happened, that proved stable, and that now canalize everything that happens around them.
Most constraints in an introductory physics course are what engineers call “holonomic.” You can write them as fixed equations and substitute them straight into the equations of motion. The constraint disappears as an independent thing. It becomes just another term in the dynamics.
Life depends on a different kind. Pattee, borrowing from analytical mechanics, called them non‑integrable constraints. Imagine a coin rolling across a table without slipping. The “no‑slip” condition ties its forward motion to its rotation, yet you cannot integrate that condition away. The coin can still reach any point on the table, but it must follow specific curved paths to get there. The constraint shapes every trajectory while leaving the full space of possibilities intact.
A ribosome sitting on a messenger RNA is a non‑integrable constraint. It couples the sequence of codons to the sequence of amino acids. It “reads” the memory, physically, without consuming it. The same mRNA can be read again and again. If the coupling were integrable, the memory would collapse into the dynamics, the correspondence between sequence and function would become energetically forced, and the capacity to store arbitrary information would vanish. The constraint is what keeps the symbol and the process distinct.
The Epistemic Cut Is Not a Wall
This coupling of a quiescent memory to a rate‑dependent process is what Pattee christened the epistemic cut. He was careful with the word. “Epistemic” doesn’t mean physical. It means “having to do with knowledge.” The cut is not a membrane, not a spatial boundary. It’s a functional distinction between the thing that specifies and the thing that executes.
The word “cat” is not a cat. The gene for insulin is not insulin. If you erase that distinction—if you insist on describing everything as one seamless dynamical system—the concept of reference dissolves. You no longer have a message, a code, or a controller. You just have particles.
John von Neumann proved this in the abstract for quantum measurement. Treat a measuring device and the system it measures as a single combined dynamical system, and the device ceases to be a measurer. It becomes just another heap of particles. To regain the measurement function, you need a new observer outside the combined system—and then another, in an infinite regress. Pattee saw that biology embodies the same logic. Describe an enzyme atom‑by‑atom, tracing every electron, solving its full quantum dynamics, and at that level of description the enzyme is no longer an enzyme in any functional sense. It is a particular arrangement of atoms. Catalytic specificity, coding, error—all of it vanishes into the equations.
This does not mean life has non‑physical ingredients. It means that functional description and dynamical description are two irreducibly different ways of carving the same reality. You can flip between them. You cannot merge them without losing something. The epistemic cut is not a flaw in physics. It is a necessary feature of any description that includes signs.
The Four‑Part Engine
Pattee’s architecture, as it crystallized in his 2001 paper “The Physics of Symbols: Bridging the Epistemic Cut,” rests on four interlocking ideas. Together they form a checklist for what any material system must have to be alive rather than merely complex.
Energy degeneracy. The bases in DNA can be arranged in any order along the backbone without significantly changing the molecule’s overall energy. A gene for a digestive enzyme and a gene for a neurotransmitter receptor have almost identical masses and thermal stabilities. Physics is indifferent to the sequence, and that indifference is everything. It means history can record itself, because no sequence is thermodynamically favored. Without degeneracy, arbitrary memory is impossible.
Non‑integrable constraints—the reading hardware. The ribosome, the spliceosome, the transcription apparatus. These structures couple a symbolic sequence to a dynamical process while keeping the two logically distinct. The mRNA is not altered by being read. The constraint steers the chemistry without being chemically changed by it.
The epistemic cut itself. The functional boundary that separates symbol from referent, instruction from execution. It is not a physical wall; the same object can sit on both sides depending on context. A stretch of RNA is a message when read by the ribosome, and a mere nutrient when digested by a nuclease. The cut is in the relationship.
Semantic closure. The machinery that reads the genetic code—ribosomes, tRNA synthetases, polymerases—is itself made of proteins specified by that same code. This is not a vicious circle. It’s a loop that can sustain itself, replicate, and vary. The code builds its own interpreters. The interpreters execute the code.
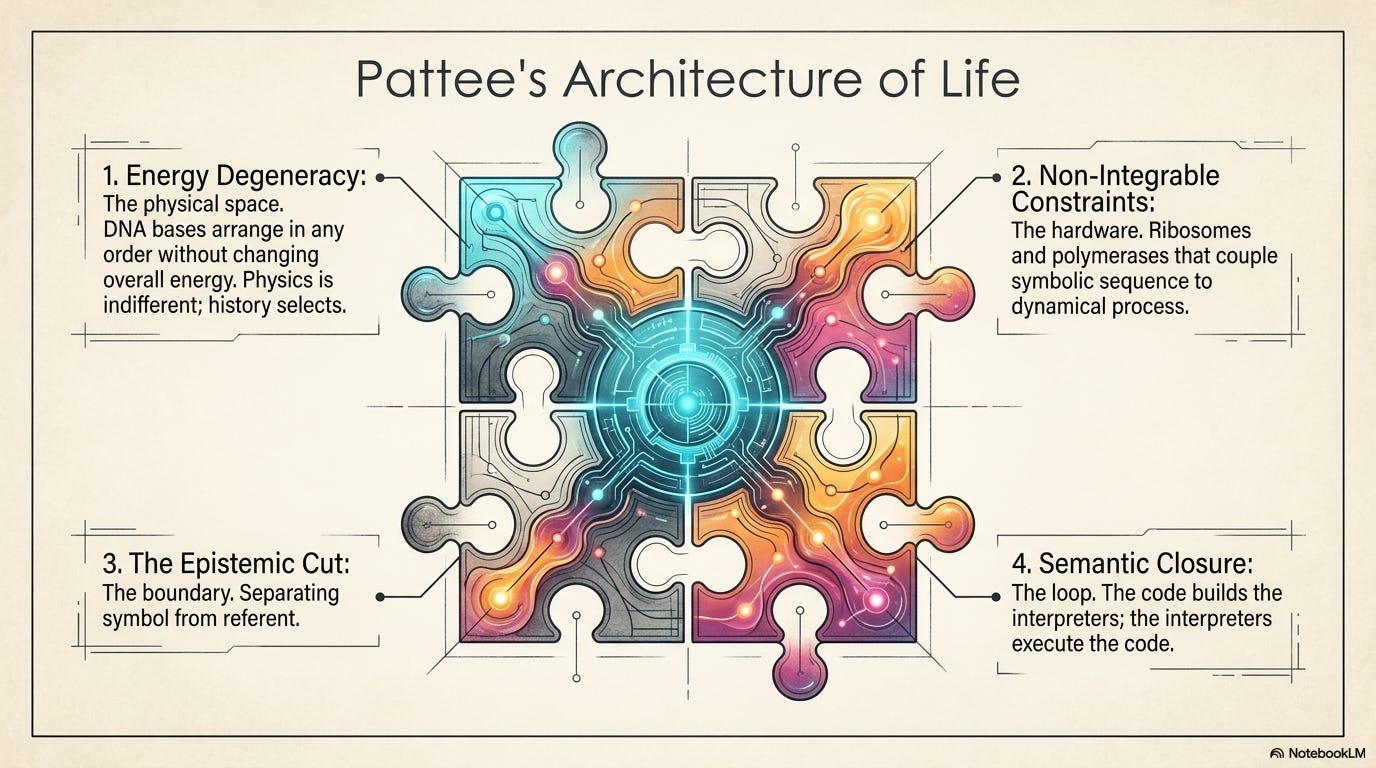
These four conditions are necessary. Pattee was explicit that they are not sufficient. They don’t tell you how the first non‑integrable constraints were assembled, or how degeneracy became coded rather than noise. But they tell you what to look for. They turn the old question—what distinguishes the living from the lifeless?—into a set of physical, testable criteria.
After Pattee: Barbieri, Hoffmeyer, Deacon
Before I bring Pattee’s framework back to condensates, I want to acknowledge three thinkers who have pulled his architecture in different directions, because doing so will clarify what my earlier essay was actually trying to do.
Marcello Barbieri formalized Pattee’s insight into a theory of organic codes. He argued that a genuine biological code requires three things: independent sequences on both sides of the mapping, molecular adaptors that enforce the mapping, and arbitrariness—meaning the mapping is not dictated by chemical affinity. The genetic code is the prototype, but Barbieri showed that splicing codes, signal transduction codes, and histone codes follow the same logic. His contribution was to treat codes not as metaphors but as literal physical artifacts—molecular adaptors that are themselves non‑integrable constraints.
Jesper Hoffmeyer took a different path. He asked what evolution does with the cut once it exists, and his answer was semiotic freedom—the expanding capacity of organisms to interpret their environments as signs rather than merely react to physical gradients. A bacterium swimming toward glucose is not being pushed; it’s reading a concentration difference as a sign, and it reads that same difference differently depending on context. Hoffmeyer argued that this interpretive flexibility is what evolution elaborates over deep time, from minimal bacterial sign‑relations all the way up to human culture.
Terrence Deacon tackled the problem Pattee left open: how semantic closure bootstraps from pure chemistry without presupposing the very symbols it is supposed to explain. Deacon’s notion of absential causation—causation organized around what is not physically present—is a genuinely subtle idea. A seed grows toward a mature form that doesn’t yet exist. Homeostasis maintains a state defined by the absence of deviation. Deacon’s autogen model demonstrates, in principle, how a minimal network of mutually catalytic molecules could cross the threshold into self‑reference, generating a functional inside‑outside distinction without any pre‑existing code. The first sign was not a genetic message. It was a molecular configuration that made a functional difference to a self‑maintaining network.
These three thinkers completed different pieces of Pattee’s map. Barbieri formalized the physical structure of codes. Hoffmeyer traced the evolutionary logic of sign use. Deacon proposed a thermodynamic pathway into the loop. But what none of them did—and what my condensates argument tries to do—is identify a concrete material entity in which the epistemic cut is instantiated within a single physical object, where the symbolic and the dynamical are not parsed into separate molecules but operate simultaneously in the same assembly.
Life Is Interpretation—What the Condensates Add
Read through Pattee, the condensate claim becomes more precise. A condensate is a non‑integrable constraint made of hundreds of weak, transient bonds. It is energy‑degenerate: hundreds of different molecular compositions, with different functional outcomes, are accessible at nearly identical free energies. It instantiates the epistemic cut internally: the same droplet carries the eRNA “text,” concentrates the reading machinery, and rewrites the chromatin state. It participates in semantic closure: the condensate assembles from proteins whose genes were activated by a previous round of condensate‑mediated reading. The architecture is Pattee’s. But the material form is new.
Pattee’s paradigm case, the ribosome, is a fixed machine enforcing a universal code. The same codon means the same amino acid in every cell, every organism, four billion years running. A condensate is something structurally different: a local, on‑demand interpreter. It assembles differently in a stem cell and in a committed neuron. It responds to developmental context, metabolic state, stress. It can dissolve in response to a signal and reform with an altered composition that shifts how the next reading proceeds. The condensate doesn’t merely instantiate the epistemic cut—it makes the cut context‑sensitive and revisable, not once in evolutionary time but continuously within a single cell’s lifespan.
This is where the “life is interpretation” thesis extends Pattee’s architecture in three directions Pattee himself left open.
The cut must be actively maintained, not passively possessed. Pattee identified the epistemic cut as what a living system has—the structural feature that distinguishes it from mere chemistry. The condensate picture insists it is something the cell does. With each write‑read‑rewrite cycle, the functional distinction between the symbolic and the dynamical must be regenerated. If the loop slows, the condensate destabilizes; symbol and process collapse back into undifferentiated chemistry. Interpretation is not a stable property sitting in a cell like a component. It is a sustained activity—perpetually reconstructed by the very process it makes possible. This is what endogenous unfreedom means: the constraint is not imposed from outside; it is generated, cycle by cycle, from within the system’s own operation. The living cell is not free to stop reading without ceasing to be a cell.
History is constitutive of the reading side of the cut. Pattee’s non‑integrable constraint reads a symbol. But the condensate reads a symbol in the context of what every previous reading has written into the system’s material state. The same enhancer RNA in a stem cell and in a committed neural precursor activates different genes—not because the sign changed, but because the reader did. The chromatin’s current tag pattern is the residue of every developmental signal the cell has encountered. The reader is its accumulated history, materially encoded. There is no substrate reader underneath that history to which the history is then added. This makes interpretation irreducibly diachronic in a way that distinguishes it from signal transduction: transduction can be synchronic, a one‑to‑one input‑output mapping. Interpretation cannot. The response is always a function of what has already been read.
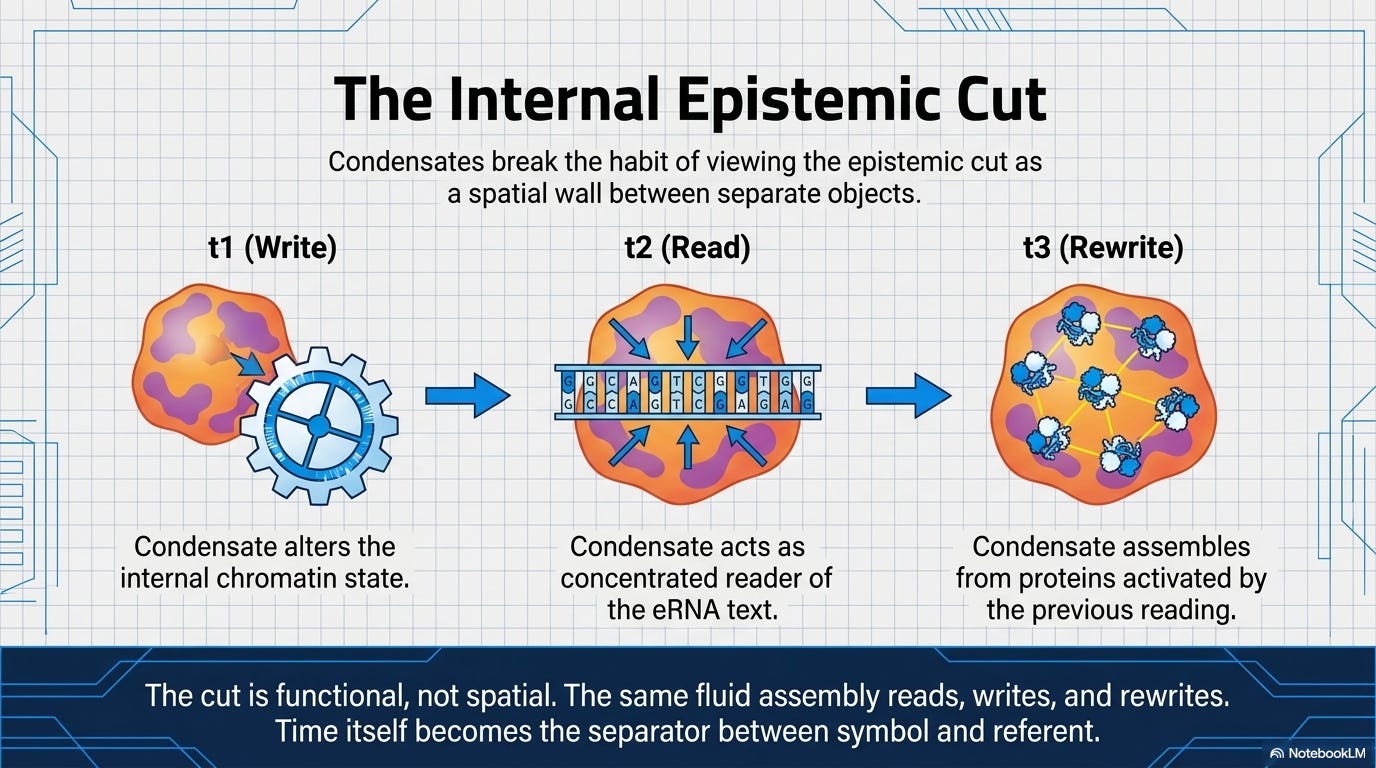
Error is the engine of the cut’s elaboration, not a deviation from correct operation. For Pattee, semantic closure stabilizes the system; error threatens it. The condensate picture inverts this. A condensate that misreads an enhancer RNA, writes a histone mark in the wrong location, or stabilizes a slightly altered composition is not malfunctioning. Most such misreadings are neutral or erased. But a productive misreading—one that opens a new chromatin region, alters a cell’s trajectory, or in evolutionary time gets fixed—is simultaneously the first instance of a new sign. The cut doesn’t merely tolerate error. It develops through error. What Hoffmeyer called semiotic freedom—the capacity to interpret the environment rather than merely react to it—accumulates exactly this way: not through design but through the selective retention of interpretive mistakes. Every retained productive misreading expanded the range of what the system could subsequently mean. A frozen accident (Gell-Mann’s) is just a productive error that selection decided to keep. The condensate is where that decision is still being made.
The condensate makes all three properties visible in a single molecular object: a liquid process whose momentary configuration encodes the history of every previous reading; whose lability is precisely what allows interpretation to be local and context‑specific rather than universal and fixed; and whose productive fallibility is what has been elaborating the epistemic cut for four billion years. That is what it looks like when matter organizes itself not merely to obey signs but to generate new ones.
Conclusion: The Physicist Who Drew the Map
Howard Pattee never solved the origin of life. He never built a living system in a test tube. But he did something harder: he stopped the question from collapsing into mysticism on one side or reductionism on the other. He said, here is the architecture. These are the necessary physical conditions. Now go look.
He considered himself a physicist, not a biosemiotician—he wanted to know how you build a sign from molecules, not how you interpret molecules as signs.
Pattee drew the map. He gave us the necessary architecture: energy degeneracy, non‑integrable constraints, the epistemic cut, semantic closure. Without that map, we would still be waving our hands about “information” and “codes” without knowing what physical conditions make them possible.
But the thesis that life is interpretation—not just that life uses interpretation, but that interpretation is the specific material activity that distinguishes living from non‑living matter—goes further. It insists that interpretation is irreducibly temporal, historical, and error‑laden. The write‑read‑rewrite loop is not an extra feature; it is the form semantic closure takes when it actually runs. History is not stored beside the reader; the reader is its own accumulated history, written into chromatin, condensate composition, and the architecture of every molecular machine shaped by past readings. Error is not a malfunction of the cut; it is the engine that has been elaborating the cut for four billion years.
The condensates make all of this visible at the molecular level—a liquid droplet that reads, writes, and rewrites; that holds its own history in its composition; and that generates novelty through the very imperfection of its readings.
Pattee answered the question Pearson asked: what distinguishes the living from the lifeless? The architecture of interpretation. The condensates show us what that architecture looks like when it’s still liquid, still becoming. The question from here is not whether matter can interpret. It’s why we ever thought it couldn’t.
References
Pattee, H.H. (2001). “The Physics of Symbols: Bridging the Epistemic Cut.” Biosystems 60: 5–21. (The single clearest statement of the four‑part architecture.)
Deacon, T.W. (2012). Incomplete Nature: How Mind Emerged from Matter. W.W. Norton. (The most sustained attempt to bootstrap semantic closure from thermodynamic processes.)
Hoffmeyer, J. (2008). Biosemiotics: An Examination into the Signs of Life and the Life of Signs. University of Scranton Press. (The evolutionary logic of semiotic freedom.)
Barbieri, M. (2003). The Organic Codes: An Introduction to Semantic Biology. Cambridge University Press. (The formal theory of biological codes as physical artifacts.)
Newman, S.A. & Sarkar, S. (2025). “Biology and Physics.” Encyclopedia of Philosophy of Science. (The argument that condensates represent a new form of matter.)
It's worth noting that there will be a symposium in October to celebrate Howard Pattee's 100th birthday. It will feature talks by many of his former students and collaborators. The date is October 15th and the venue is Binghamton University, where he taught for many years.