
Life Is Interpretation
Biomolecular Condensates, the Epistemic Cut, and Interpretation as a Form of Matter

I. The Discovery That Changes the Question
Inside every living cell, something is happening that contemporary physics cannot explain — not because the details are complicated, but because the kind of thing happening does not fit within any existing physical framework.
The cell’s interior is not a uniform soup. It is organized into compartments, specialized regions that concentrate particular molecules and carry out particular tasks. Most of these compartments are surrounded by membranes — thin fatty barriers that physically wall off their contents from the rest of the cell. Mitochondria, the endoplasmic reticulum, the Golgi apparatus: these are membrane-bound organelles, and their containment mechanism is straightforward. A physical wall keeps their contents in and everything else out.
But over the past two decades, biologists have discovered that the cell also contains dozens of compartments that have no membrane at all. These are biomolecular condensates (BMCs) — concentrated assemblies of proteins and RNA molecules that form coherent, bounded bodies within the cell, maintain their identity, carry out specific functions, and then dissolve when their work is done, all without any enclosing wall. They are held together not by a container but by the collective stickiness of their molecular components: thousands of weak, transient, reversible interactions, each lasting only milliseconds, but whose collective effect produces a structure that persists for minutes to hours.
Stuart Newman and Sahotra Sarkar, in a recent paper, Biology and Physics (2025), argue that BMCs represent a genuinely new form of matter — a kind of material organization whose properties cannot be derived from any existing physical theory. This is a strong claim. Understanding why they make it, and what it implies, requires examining what BMCs actually do — and then asking a question the paper itself does not fully pursue: whether interpretation, the activity BMCs perform, might itself constitute a new form of matter.
II. What BMCs Do
BMCs appear throughout the cell. In the cytoplasm, stress granules form when the cell is threatened — they sequester messenger RNAs and pause non-essential protein production while the cell manages the emergency. When the crisis passes, the granules dissolve and normal operations resume. In the nucleus, the nucleolus — the largest structure in the nucleus — is a BMC where ribosomes are manufactured. It has its own internal architecture, concentric layers with different compositions and functions, all maintained without any internal membranes.
Most consequential for the argument that follows are transcriptional condensates, which Newman calls function-amplifying centers (FACs). These are nuclear BMCs that form at specific locations on the genome and coordinate the activation of specific genes. They are the structures most directly involved in determining what kind of cell a cell becomes — whether it develops into a neuron, a muscle cell, a liver cell, or any of the roughly 300 distinct cell types in the human body.
The mechanism works as follows. Enhancers — short regulatory DNA sequences scattered throughout the genome — are transcribed into small RNA molecules called enhancer RNAs (eRNAs). These eRNAs nucleate condensate formation at specific genomic locations, recruiting transcription factors and the Mediator complex (a large protein assembly that bridges regulatory proteins to the gene’s transcription machinery). The condensate concentrates this machinery at specific genomic sites. Through chromosomal looping, these sites are brought into physical contact with target gene promoters. The result is intense, coordinated transcription of a specific set of genes — which is to say, a specific cell type.
Which genes get activated depends on which eRNAs are present, which depends on cellular history and context. The same genome, read through different condensate configurations in different contexts, produces different cell types. The genome is not a program being executed. It is a text being interpreted.
III. Five Properties, One Entanglement
Newman and Sarkar’s central claim is that BMCs combine five distinct functional properties simultaneously, and that the combination — not any single property — defeats existing physical theory. Each property, taken alone, might be accommodated by extending known frameworks. Their simultaneous presence and mutual dependence cannot.
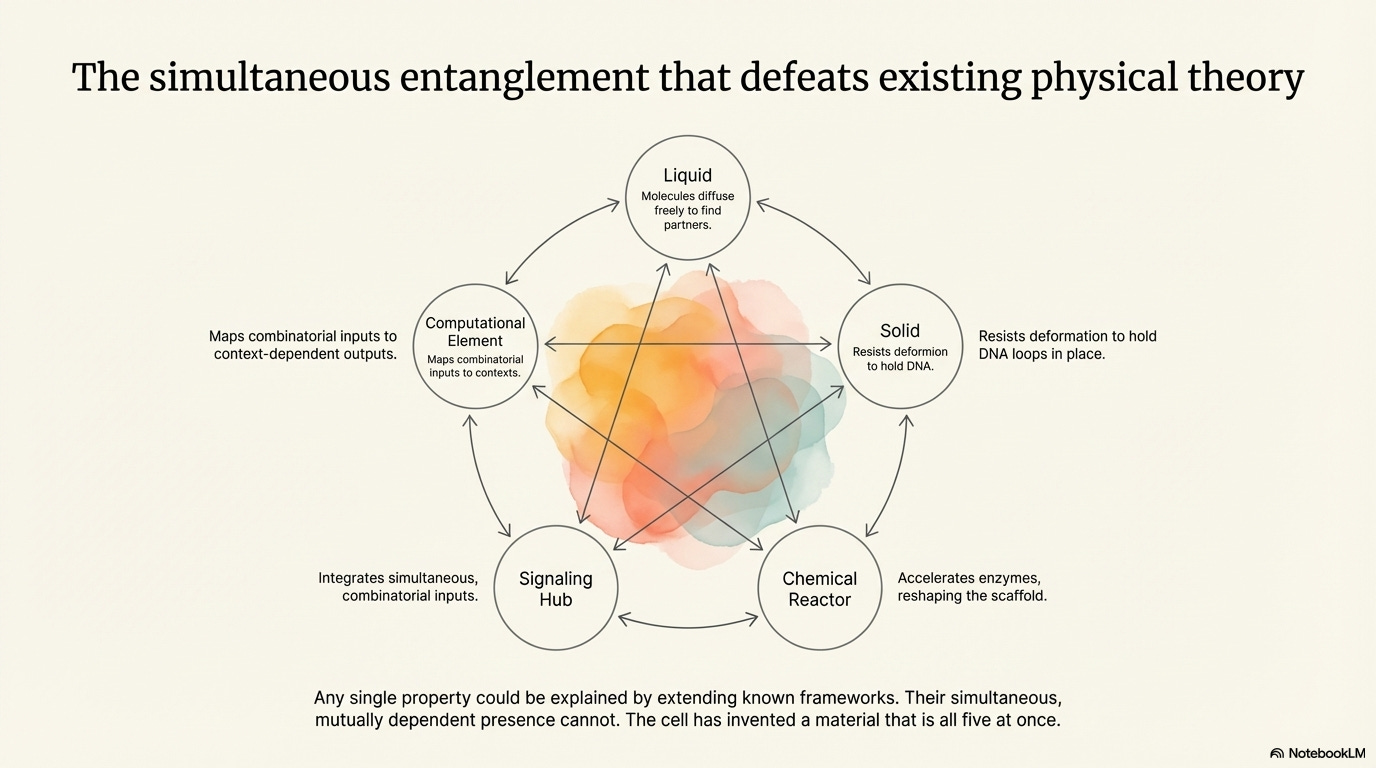
A Liquid
The molecules inside a BMC are in constant motion, diffusing freely, rearranging, finding new partners. Two BMCs of the same type that come into contact will merge, flowing into each other like water droplets on glass. Classical liquid theory is powerful but built for systems of interchangeable particles. In a BMC, the ‘liquid’ contains dozens to hundreds of chemically distinct molecular species, each with specific partners and specific roles. The specificity of the components is essential to the condensate’s function. No existing liquid theory accommodates this combination of fluidity and molecular specificity.
A Solid
At certain timescales, the same BMC resists deformation like a solid — holding chromosomal loops in position, maintaining the three-dimensional genomic architecture required for coordinated gene activation. This is not the rigidity of a crystal (there is no repeating lattice) nor the kinetic trapping of a glass (BMCs dissolve on demand). Their mechanical properties are actively maintained by continuous energy consumption and can be tuned up or down in response to specific molecular signals. No existing theory of solids describes a material whose rigidity requires continuous energy input and is dynamically adjustable.
A Chemical Reactor
BMCs concentrate specific enzymes and substrates, accelerating reactions sometimes by orders of magnitude compared to the surrounding cell. But unlike any reactor in chemical engineering, the BMC has no fixed walls. Its boundary is a concentration gradient. Its composition changes continuously as molecules diffuse in and out. Most significantly, the reactor is itself made of molecules that are simultaneously reactants and products of the reactions occurring within it. The vessel is produced by, and continuously reshaped by, its own chemistry.
A Signaling Hub
A transcriptional condensate does not merely accelerate a single reaction. It integrates multiple molecular signals arriving simultaneously — enhancer RNAs from different genomic regions, transcription factors carrying signals from outside the cell, chromatin state information, mechanical signals from the cell’s structural skeleton — and produces a combinatorially specific output: the activation of a particular set of genes at particular levels. Existing signaling theory models molecular communication as chains of discrete events. In the BMC, signal processing is simultaneous and compositional. The ‘signal’ is not a molecular event but the overall material composition of the condensate at a given moment.
A Computational Element
The function-amplifying center receives a combinatorial input — which enhancer RNAs are present, in what relative abundances — and maps it to a specific transcriptional output. The mapping is context-dependent: the same eRNA combination in a cell with a different developmental history produces a different output. This is computation in a non-trivial sense, but unlike any computation existing theory describes. There are no discrete logical gates. No identifiable wiring connects inputs to outputs. The computation is a property of the condensed material as a whole.
The Entanglement
These five properties are not independent features that happen to coexist. They are mutually constitutive — each depends on the others, and removing any one destroys the system’s function.
The liquid character enables the reactor and signaling functions: molecules must diffuse freely to find each other and integrate. The solid character enables the computational function: mechanical stability holds chromatin loops in position long enough for specific gene activation. The reactor character maintains the liquid and solid properties simultaneously: enzymatic reactions continuously modify scaffold proteins, dynamically tuning the condensate’s mechanical character. The signaling character provides the context-dependence that makes the computation biologically meaningful. The computational character closes the loop: newly activated genes produce new eRNAs that feed back into the condensate, modifying its composition and its next round of output.
This is what Newman and Sarkar mean when they call BMCs a genuinely new form of matter: not that any single property is unprecedented, but that their mutual entanglement produces something for which existing physical frameworks provide no starting vocabulary. The cell has, in effect, invented a material that is simultaneously a liquid, a solid, a chemical reactor, a signaling hub, and a computational element — and we have almost no physical theory that captures all of these properties at once.
IV. The Epistemic Cut Runs Through the Condensate
To understand why BMCs are philosophically significant — and not merely physically puzzling — we need a concept developed by the physicist and theoretical biologist Howard Pattee, who spent his career asking what distinguishes living matter from nonliving matter. His answer centers on what he called the epistemic cut: the fundamental distinction between physical dynamics and the symbols that describe or control them.
Pattee’s key observation is that every living system contains two functionally distinct kinds of component. On one side: structures that function as symbols — whose causal power depends not on their physical properties but on their specific configuration. DNA is the paradigm case. A stretch of DNA encoding one protein has nearly identical physical properties to a stretch encoding a different protein. What matters is the sequence — the specific order of chemical letters — not the molecule’s mass, charge, or reaction rates. The sequence instructs the cell’s machinery, but the sequence itself does not push, pull, or catalyze anything.
On the other side: structures that do the dynamical work — the proteins, molecular machines, and enzymes whose causal power depends on their physical and chemical properties. These read the symbolic side and act on it.
The deepest feature of the cut: neither side can be derived from the other. No amount of physical analysis of DNA will tell you what it means, what proteins it specifies, what the cell will do with it. The meaning is constituted by the reading machinery, not by the DNA’s physics. The mapping between symbol and action is logically arbitrary relative to physics — there is no physical law that requires a particular genetic sequence to specify a particular amino acid. The code is a convention, maintained by evolved machinery rather than natural necessity. Alternative codes are physically possible. This is what Pattee means when he writes that the genetic code exemplifies ‘rate-independent constraints on rate-dependent processes’ — the symbol constrains the dynamics without being determined by them.
In the standard picture of molecular biology, the two sides of the cut are carried by physically distinct objects. You can extract DNA and study it in isolation. You can purify proteins and study them separately. The symbolic and dynamical functions are parceled out to different molecular species, which is part of why molecular biology has been so successful.
BMCs violate this clean separation. The condensate is simultaneously the thing that carries the meaningful configuration — which eRNAs, which transcription factors, which combination specifying which genes — and the thing that reads and acts on that configuration. It concentrates the enzymes, catalyzes the reactions, rewrites the chromatin. It is both the text and the reader. The epistemic cut runs not between objects but through a single material structure.
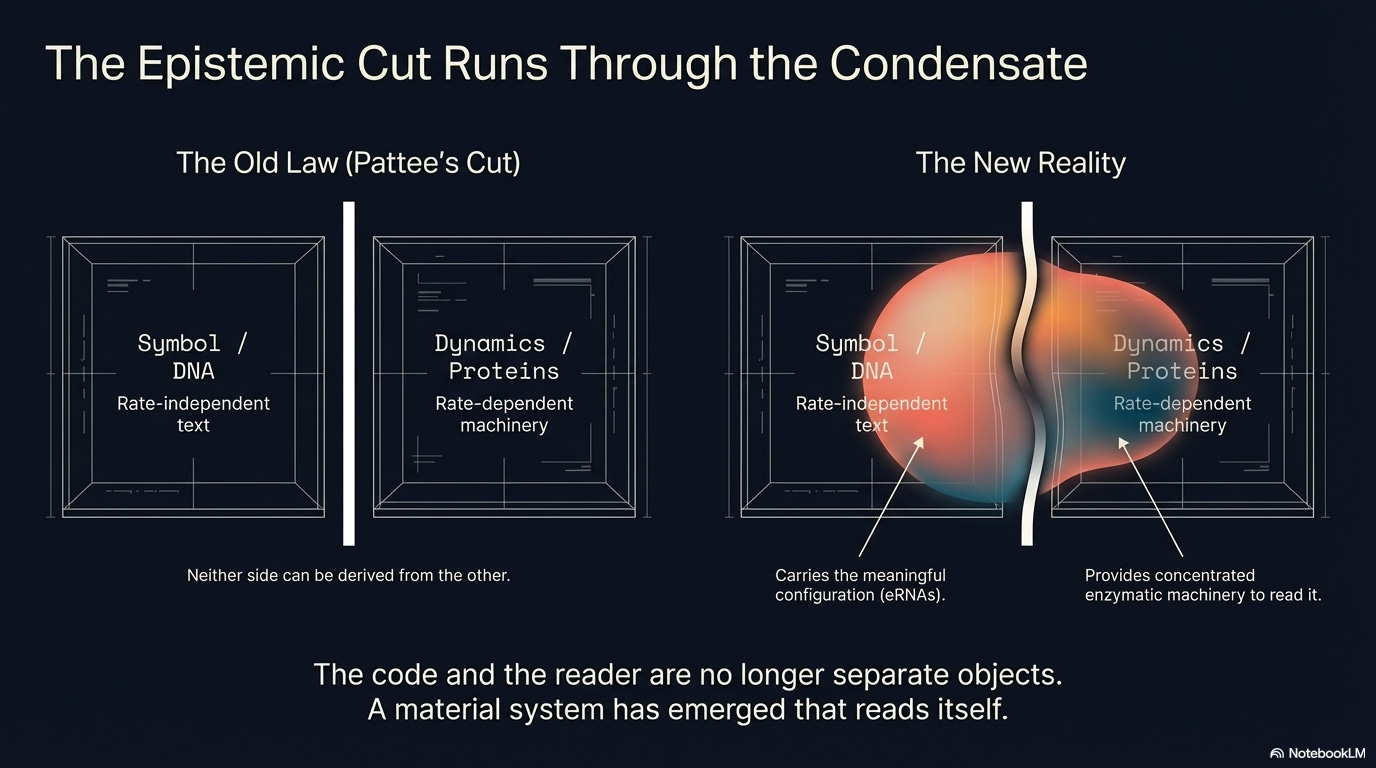
This is not a contradiction of Pattee’s framework. It is a revelation of its deeper structure. The cut is functional, not physical. The same object can operate in both modes simultaneously. And when it does, something new is possible: a material system that reads itself.
V. The Write-Read-Rewrite Loop
The biological function that makes this most consequential is cell differentiation — how a single genome gives rise to hundreds of distinct cell types.
DNA in the cell is wound around spool-like nucleosome complexes of histone proteins, whose flexible tails can be chemically tagged with small groups — methyl, acetyl, phosphate — at specific positions. These tags constitute a code. Certain combinations mark a genomic region as active, others as silenced, others as poised for activation. The tags are written by enzymes that attach specific groups, read by proteins with specialized recognition domains, and rewritten by enzymes that remove existing tags and replace them with different ones. This is what Newman calls the write-read-rewrite system: a form of molecular memory that is durable but not permanent, specific but not fixed, and self-modifying.
The self-modification is the crucial feature. When a cell receives a developmental signal, activated transcription factors nucleate BMC assembly at specific enhancer loci. The condensates recruit histone-modifying enzymes that rewrite the chemical tags on nearby nucleosomes, opening some genomic regions and closing others. The newly opened regions include enhancers that produce new eRNAs. These new eRNAs modify the condensates, which change the pattern of gene activation further. The newly activated genes include both structural proteins that define the cell type and regulatory factors that stabilize the new identity — including new eRNAs that feed back into the condensates again.
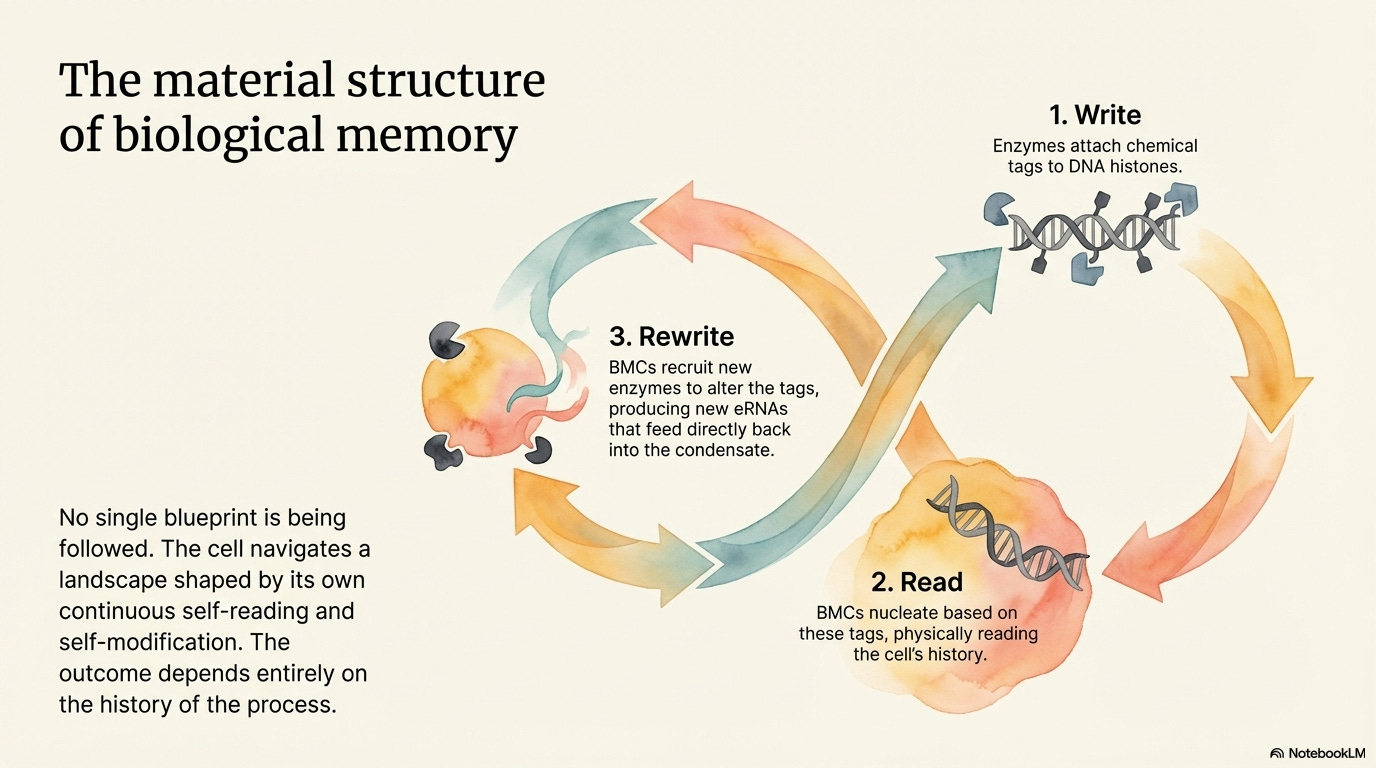
The system reads its own state, writes new marks, reads those marks, and rewrites again. No single program is being executed. The cell is navigating a landscape shaped by its own continuous self-reading and self-modification. The outcome depends not on a blueprint but on the history of the process — which eRNAs were present at which moment, which modifications accumulated in which order, which signals arrived when. History is not stored separately from the system. It is written into the material state of the chromatin, held in the condensates, expressed through the current round of reading.
This is the write-read-rewrite loop: the material structure of interpretation at the molecular level.
VI. Life Is Interpretation
The write-read-rewrite loop is not a feature peculiar to cell differentiation. It is the organizational principle of living systems at every scale. This is the claim of biosemiotics — and the evidence for it, examined carefully, goes further than analogy.
A bacterium swimming toward a sugar gradient is not merely executing a chemical response. Its behavior depends on context: the same sugar signal is read differently depending on what other nutrients are present, what the bacterium has already consumed, and what threats surround it. A thermostat responds to temperature the same way regardless of any other variable. The bacterium’s response to any given signal depends on what else is present — and that context-dependence is itself shaped by selection to produce adaptive outcomes across varied situations. This is not mere mechanism. It is interpretation in a precise sense: a functional reading of signs in context, where the reading apparatus was built by the same selective process that shaped the text.
The immune system reads molecular signatures on cell surfaces and interprets them as self or non-self — and the interpretation is context-dependent, because the same molecular pattern that triggers an immune attack in one tissue is tolerated in another. Plants interpret caterpillar damage by releasing airborne chemicals that attract parasitic wasps — a sign emitted by one organism, read by another, to the detriment of a third. The same gene, Pax6, initiates eye development in both mice and flies, producing a camera eye in one and a compound eye in the other. Same text, radically different interpretation, both functional. Testosterone promotes aggression in some social settings and paternal care in others. Same signal, different reading, different outcome — and the context-dependence is itself evolved.
What Jesper Hoffmeyer called semiotic freedom — the increasing capacity of organisms to interpret their environments flexibly — runs continuously from bacterium to philosopher. The basic structure is the same at every level: sign, interpretation, context-dependent response, history-dependent reading. The difference between a bacterium and a human is a difference in degree of semiotic complexity, not a difference in kind. Interpretation is not something that appears when nervous systems reach a certain size. It is the organizational principle of life from the beginning.
This is the commitment the BMC evidence forces: life is interpretation. Not that living systems can be described in semiotic terms as a useful analogy. Not that interpretation is an emergent property of sufficiently complex organisms. But that the structural logic of interpretation — sign, reading, context, history, error, coherence, self-modification — is what distinguishes living matter from nonliving matter at every scale where the distinction holds.
VII. Interpretation as a Form of Matter
If life is interpretation, then the philosophical question raised by BMCs is not merely ‘what kind of material are these?’ It is: what kind of thing is interpretation itself?
Newman and Sarkar work in the tradition of dialectical materialism, which holds that matter exists in qualitatively distinct forms — levels of organization, each with its own characteristic mode of activity and its own laws irreducible to the laws of lower levels. Friedrich Engels identified a hierarchy: mechanical motion, physical motion, chemical motion, biological motion, social motion. The word ‘motion’ is technical — it means not locomotion but the characteristic activity of each level. The laws of chemistry cannot be derived from the laws of mechanics, even though every chemical reaction involves mechanical motion of atoms. The laws of biology cannot be derived from the laws of chemistry, even though every biological process involves chemical reactions. At each level, genuinely new regularities appear that belong to that level and to no lower level.
Newman and Sarkar’s ‘inherencies’ — the dispositional tendencies built into each form of matter — are this framework applied to biological materials. Liquid tissue has the inherency to minimize surface area, to phase-separate by adhesion, to flow. These are not imposed from outside. They are what that kind of matter does.
The question is whether interpretation satisfies the structural requirements for a form of matter in this sense. There are five such requirements, and interpretation meets each of them.
Materiality. Every instance of interpretation that biosemiotics describes is materially implemented. The first productive error — a copying mistake in a self-replicating molecule that happened to work and was retained by selection — was a molecular event. Cellular reading happens through BMCs, physical assemblies of proteins and RNA obeying the laws of chemistry. Neural interpretation happens through electrochemical processes. Human interpretation happens through brains, vocal tracts, written texts, institutional structures. At no point is an immaterial substance required. Meaning is not a ghost. It is what certain material organizations do.
Qualitative novelty. Before the first productive error, molecules reacted. Their behavior was determined by their physical and chemical properties. There was no gap between what a molecule is and what it does. After the first productive error, a new kind of activity appeared: a system whose behavior depends not only on its current physical state but on its accumulated history of readings — on the products of previous rounds of copying, variation, selection, and reinterpretation. A cell’s chromatin state means something different — activates different genes, specifies a different cell type — depending on the sequence of readings that produced it. This history-dependent, context-sensitive, self-modifying activity is absent from chemistry. It is a qualitative novelty.
Own laws. The interpretive loop — error generates variation, resistance tests it, coherence selects what works, successful readings are externalized as new structure for future readers, imperfect future readings generate new variation — is a structural regularity that operates wherever interpretation occurs: in molecular evolution, in cellular differentiation, in neural learning, in cultural transmission. This regularity involves categories — sign, reading, context, history, coherence — that have no counterparts in physical or chemical theory. The context-dependence of an eRNA in a BMC is not a chemical law. No chemical law relates molecular properties to outcomes with reference to interpretive history. The regularity belongs to the interpretive level and to no lower level.
Historical emergence. Interpretation emerged from chemistry through a specific, natural, material event: the first productive error. It did not exist before that event. It came into being through molecular accident and has been developing ever since through further material processes: the evolution of cells, nervous systems, language, culture. Its genealogy is continuous and material.
Characteristic mode of activity. Interpretation’s characteristic activity is reading: responding to specific configurations of matter not merely according to those configurations’ physical properties but according to what those configurations specify in the context of the system’s accumulated history. Reading is to interpretation what metabolism is to life: the activity that defines the form.
Interpretation satisfies all five requirements. It is fully material, qualitatively novel, governed by its own irreducible regularities, historically emerged, and characterized by a distinctive mode of activity. It is a form of matter.
VIII. Dissolving the False Dilemma
If interpretation is a form of matter, the long-standing oscillation between physicalism and dualism is not just resolved — it is dissolved. The choice was never between ‘everything is physics’ and ‘meaning requires something non-material.’ There is a third option, and it is the one the BMC evidence most naturally supports: some things are not physical but are still material. They belong to higher forms of matter with their own irreducible properties.

Meaning is not a property of matter in general. It is a property of matter organized at the interpretive level — matter that carries the write-read-rewrite structure, that sustains the epistemic cut within a single physical object, that accumulates history as functional constraint on future readings. Not matter plus meaning. Not matter decorated with signs. Matter as sign — organized in the specific way required for interpretation to occur.
This reframing has consequences for every framework in this discussion.
For Newman and Sarkar’s project: the search for a theory of BMCs would be reframed. The goal would not be to extend condensed matter physics to cover BMC properties — which they have shown cannot work — nor to wait for new physics to appear from nowhere. It would be to develop a theory at the interpretive level: a theory whose primary categories are signs, readings, contexts, histories, errors, and coherences, and whose regularities describe the structural logic of interpretive processes. This theory would be as materialist as thermodynamics but irreducible to it, just as thermodynamics is materialist but irreducible to Newtonian mechanics.
For biosemiotics: the framework gains a materialist grounding it has not always been able to claim clearly. Meaning is not something beyond the material — a characterization that, however carefully qualified, invites accusations of dualism. It is something within the material at a specific level of organization. The interpretive gap is not a gap between the material and the non-material. It is a gap within the material — the space of irreducibility between lower forms (physics, chemistry) and the interpretive form. The same kind of gap that exists between mechanics and chemistry, or chemistry and biology. Real, permanent, irreducible, and entirely within the material world.
For the origin of life: the question shifts from ‘what was the first self-replicating mechanism?’ to ‘what was the first self-reading structure?’ The origin of life becomes the origin of interpretation — the first moment when a material system’s behavior depended not only on its current physical state but on what that state specified for a reading process. The first productive error is not merely the first mutation. It is the first sign.
IX. The Stakes
If interpretation is a form of matter with its own irreducible laws, this is not a philosophical conclusion that leaves biology unchanged. It carries specific scientific consequences.
It means that a complete physical description of a BMC at a given instant does not capture what the condensate means — which cell type it specifies, how the current reading relates to the history of previous readings, what future readings it makes possible. Meaning is a relational, historical, context-dependent property that no instantaneous physical description can encode. The explanatory gap between BMC physics and cell identity is therefore not temporary — not a gap that better physics will close. It is structural. It belongs to the relationship between forms, not to the limits of current theory.
It means that the categories of molecular biology — program, blueprint, code, information — are not wrong but are systematically incomplete. They describe the symbolic side of the epistemic cut while leaving the reading process untheorized. A genetics that describes genes without describing the interpretive machinery that reads them, in context, with history, is a genetics that has not yet accounted for what genes actually do.
It means that developmental system drift — the evolutionary phenomenon whereby the same morphological outcome comes to be produced by different underlying mechanisms — is not merely evolutionary noise or robustness-seeking. It is the gradual installation of a semiotic layer on top of a physical substrate: the replacement of a direct physical process by a genetic machinery that reads a signal to produce the same outcome. Each step in this replacement is a step toward greater semiotic freedom — greater context-sensitivity, greater capacity for history-dependent modulation. Developmental system drift may be the evolutionary trajectory of interpretation itself.
And it means that the emergence of organisms capable of reflecting on their own interpretive processes — capable of noticing the interpretive gap, examining its material basis, and asking what it means — is not a discontinuity in nature but a continuation of the same process that began with the first productive error. We are not outside the loop looking in. We are the loop applied to itself: interpretation reading its own structure, writing new marks based on that reading, generating new variation, extending the gap that has been widening for four billion years.
X. The Open Question, Sharpened
Newman and Sarkar expect, as dialectical materialists, that new physical theory will eventually account for BMC properties — just as quantum mechanics accounted for atomic properties that classical physics could not explain. The gap is real but temporary; better science will close it.
The argument advanced here agrees that the gap is real but not that it is temporary. It is the permanent gap between forms of matter. No physics of BMCs, however complete, will describe what a condensate means — which developmental history it encodes, which cell type it specifies, how it participates in the continuous self-reading of a living system. This is not because meaning is beyond the material. It is because meaning belongs to a higher form of matter than physics describes, just as chemical bonding belongs to a higher form of matter than mechanical collision, and just as evolution belongs to a higher form than chemical reaction.
BMCs are the material structures that instantiate interpretation at the molecular level. Their five-property entanglement — liquid, solid, reactor, signaling hub, computational element, all simultaneously present and mutually dependent — is the material signature of the interpretive form: what matter looks like when it is organized at the level where reading becomes possible.
Life is what happens when matter organizes itself to read. The first cell was the first reader. Every organism since has been an elaboration of that original act: matter taking up its own structure as a sign and doing something with it, in context, with history, imperfectly, generating new variation in the reading, which becomes new material for the next reading.
The fact that we can examine this process, formalize it, and ask whether interpretation is a form of matter — is itself an instance of the interpretive loop turned on itself. We are reading the cell’s reading of its genome. In doing so, we are not stepping outside the loop. We are extending it.
The loop does not pause for our analysis. It runs through it.
References
Newman, S. A., & Sarkar, S. (2025). Biology and Physics. Encyclopedia of Philosophy of Science. [Preprint / forthcoming]. The central framework of this essay — the tripartite typology of living matter (generic, biogeneric, nongeneric), the concept of inherencies, and the proposal that biomolecular condensates constitute a genuinely new form of matter requiring new physical theory — is drawn from this paper.
Pattee, H. H. (1969). How does a molecule become a message? Developmental Biology Supplement, 3, 1–16. Pattee’s foundational paper on the origin of the genetic code as a rate-independent constraint on rate-dependent dynamics — the first systematic formulation of the problem that became the epistemic cut.
Pattee, H. H. (2001). The physics of symbols: bridging the epistemic cut. BioSystems, 60(1–3), 5–21. The clearest single exposition of the epistemic cut concept — the functional boundary between symbolic structures and the dynamical processes they control — and its implications for a physics of living systems.
Hoffmeyer, J. (2008). Biosemiotics: An Examination into the Signs of Life and the Life of Signs. University of Scranton Press. The foundational text for the claim that semiosis — sign-reading in context — is the organizational principle of life at every scale, from molecular replication to human language.
Haig, D. (2020). From Darwin to Derrida: Selfish Genes, Social Selves, and the Meaning of Life. MIT Press. The source of the productive-error framework and the argument that genuine teleology — purpose without a designer — follows from gene-centered evolutionary theory rigorously pursued.